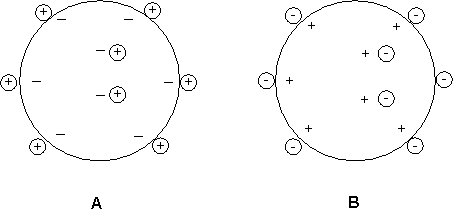
Ion-exchange chromatography
An ion-exchange resin consists of an insoluble matrix with charge groups covalently attached. Both positively charged and negatively charged ion exchangers are commercially available. Negatively charged exchangers bind positively charged ions (cations) They can bind one type of cation but, when presented with a second type of cation, this may displace, or exchange with, the first. Hence these resins are called cation-exchange resins. Similarly anion-exchange resins are positively charged and bind (and exchange) negatively charged ions (anions).
A Cation exchange resin with bound positive counterions
B Anion exchange resin with bound negative counterions
Several side-chain groups of the amino acid residues in proteins are ionizable (e.g. lysine or glutamic acid) as are the N-terminal amino and C-terminal carboxyl groups. Therefore proteins are charged molecules. This characteristic can be used to separate different proteins by ion-exchange chromatography. The two most commonly used resins for ion-exchange chromatography of proteins are carboxymethyl-cellulose (CM-cellulose) and diethylaminoethyl-cellulose (DEAE-cellulose). These are granular celluloses which have been chemically modified. CM-cellulose is a resin where the -CH2OH groups of the carbohydrate have been converted to -CH2OCH2COOH groups. At neutral pH this group is ionized as -CH2OCH2COO¯ so that CM-cellulose is negatively charged, i.e. it is a cation exchanger. DEAE-cellulose contains an ionizable tertiary amine group instead. It is positively charged at neutral pH and so DEAE-cellulose is an anion exchanger. Closely related to these ion-exchange celluloses are DEAE-Sephadex and CM-Sephadex, containing the same ionizable groups covalently attached to a Sephadex bead matrix, and the covalently cross-linked agarose bead ion exchangers DEAE-Sepharose and CM-Sepharose. DEAE-cellulose is now itself also available in a bead form called DEAE-Sephacel. The Sephadex and Sepharose types are particularly useful for the separation of high molecular weight proteins. In practice, since these matrices are very similar to those used for gel filtration some molecular sieving may accompany the ion-exchange process. This may either enhance or reduce the effectiveness of the fractionation compared to using an ion-exchange cellulose. For simplicity in the computer simulation, molecular sieving is assumed not to occur during ion-exchange chromatography. Other ion-exchange resins using different ionizable groups from those described here are also commercially available. This computer simulation includes the following ion-exchange resins: CM-cellulose, DEAE-cellulose, Q-Sepharose Fast Flow and S-Sepharose Fast Flow. The charged group of Q-Sepharose is a quarternary amine which carries a non-titratable positive charge. Therefore this matrix can be used at alkaline pH values at which the positive charge of the DEAE group would have been titrated. The charged group of S-Sepharose is the sulphonyl group (-SO3¯).
The fractionation of proteins by ion-exchange chromatography depends upon differences in the charge of different proteins. The charge of a protein depends upon the number and type of ionizable amino acid side chain groups. Lysine residues, for example, have a positively charged side chain group when ionized, whereas glutamic acid residues are negatively charged when ionized. Each ionizable side chain group has a distinct pKa; that is, the pH at which it is half dissociated. Therefore the overall number of charges on a particular protein at a particular pH will depend on the number and type of ionizable amino acid side chain groups it contains. Since, by definition, different proteins have different amino acid compositions, they will tend to have different charges at a given pH and so can be fractionated on this basis.
For any one protein there will be a pH at which the overall number of negative charges equals the number of positive charges and so it has no net charge. This is its isoelectric point (pI), or more strictly speaking its isoionic point. At this pH the protein will not bind to any ion-exchange resin. Below this pH the protein will have a net positive charge and will bind to a cation exchanger, whilst above this pH it will have a net negative charge and bind to an anion exchanger. In principle, therefore, one could choose to use either a cation exchanger or an anion exchanger to bind the protein of interest. However, proteins are usually stable (and functionally active) only within a fairly narrow pH range so that the choice of ion exchanger is often dictated by the pH stability of the desired protein. If the protein is most stable at pH values below its pI, a cation exchanger should be used whereas if it is most stable at pH values above its pI, an anion exchanger would be chosen. Clearly, if it is stable over a wide pH range, use of either type of resin can be attempted and that which gives the best fractionation is selected.
When ion-exchange resins are purchased, the ions bound to the charged groups on the resin are called 'counterions'. For CM-cellulose the counterion is usually Na+ and for DEAE-cellulose the counterion is normally Cl¯. After choice of the appropriate resin, it is mixed with buffer to form a slurry which is poured into a suitable chromatography column. The pH of this starting buffer is crucial since it will determine the charge on the proteins to be separated. The starting buffer pH should be at least one pH unit above or below the pI of the protein to be bound to the resin to ensure adequate binding. However, bear in mind that CM-cellulose and DEAE-cellulose are examples of weak ion exchangers. A weak ion exchanger is one which is ionized over only a limited pH range. Thus DEAE-cellulose begins to lose its charge above pH 9 whilst CM-cellulose begins to lose its charge below about pH 5. The term 'weak' does not refer to the strength of binding of ions to the resin nor to the physical strength of the resin itself. With these points in mind then, the effective starting pH range when using DEAE-cellulose or CM-cellulose is only about pH 5 - 9. In addition to correct choice of the pH of the starting buffer, one should take care that its ionic strength is reasonably low since the affinity of proteins for ion-exchange resins decreases as ionic strength increases. Indeed this property is used in one method of eluting the bound proteins (see below).
Once poured into the column, the bed of ion-exchange resin is washed well with starting buffer and then the protein mixture is applied. Proteins which are oppositely charged to the resin at the starting pH will bind to it, so displacing the counterions. Proteins with the same charge as the resin or with no net charge will not bind and so will flow straight through the column. The different proteins bound to the column, one of which will be the protein of interest, will have different affinities for the ion exchanger due to differences in their net charge. These affinities can be altered by varying either the pH or the ionic strength of the column buffer. Consider a set of proteins bound to the anion exchanger, DEAE-cellulose. As the pH is lowered, -COO¯ groups on the protein begin to become protonated and so lose their charge. Thus the overall negative charge of the protein will decrease and therefore so will its affinity for the resin. Different proteins will elute from the resin at different pH values as their number of negative charges decreases to a critical value. Therefore one can resolve the proteins bound to the column by slowly reducing the pH using a buffer pH gradient and collecting fractions, each of which will contain different proteins eluted at different pH values. Conversely, when using a cation exchanger, the pH gradient would be arranged to increase to elute the bound proteins.
Instead of changing pH to elute proteins bound to an ion exchanger, one can increase the ionic strength of the column buffer instead. At low ionic strength, competition between the buffer ions and proteins for charged groups on the ion exchanger is minimal and so the proteins bind strongly. However, once the proteins are bound, increasing the ionic strength increases the competition and so reduces the interaction between the ion exchanger and proteins, causing the proteins to elute. One can elute the bound proteins by increasing ionic strength irrespective of whether an anion or cation exchanger was used. This is usually accomplished by incorporating a linear concentration gradient of NaCl in the column buffer while keeping the pH constant.
Use of either a continuous pH gradient or a continuous salt (ionic strength) gradient will result in a high degree of protein fractionation based on protein charge. In practice one can also use stepwise elution, i.e. alter the pH or ionic strength in discrete known steps. Although technically simpler to carry out on occasions, stepwise gradients typically give poorer resolution than continuous gradient elution and so are not modelled in this program.